

Distribution status of seagrass and its relationship with different habitat types in Liusha bay of Guangdong province
-
摘要:
2016和2017年对广东流沙湾海草床进行现场调查,获得了海水、海草、浮游生物、底栖动物和大型底栖海藻的数据资料。结果表明流沙湾卵叶喜盐草(halophila ovalis)平均生物量为3.74 g/m2,总面积为26.9 hm2,呈斑块状分布。利用水质参数进行主成分分析,对各主成分得分进行聚类,将调查区域划分为渔排和捕鱼网影响区(Ⅰ区)、贝类养殖区(Ⅱ区)和非养殖区(Ⅲ区)3种生境类型,主成分综合得分表明Ⅰ区污染最严重。比较Ⅰ区和Ⅱ区两种不同养殖方式下的生境类型,Ⅰ区的海草生物量、底栖动物生物量、浮游植物丰度、溶解氧、活性磷酸盐和盐度低于Ⅱ区,大型底栖海藻生物量、氨氮、无机氮、N/P比值和COD高于Ⅱ区。流沙湾海草床的分布面积和生物量较2002年出现了严重退化,网箱渔排和捕鱼网的生产方式、小个体贝类的大量繁殖以及低盐度的近岸河流输入降低了海草生物量。海草生物量与其他指标的关系体现了贝类养殖对海草的生存有积极作用,较强的水体自净能力有利于海草的生长。本文还从提高公共意识、加强自然保护区管理和实施常态监测三个方面提出了流沙湾海草的保护建议。
Abstract:Field investigation was carried out in 2016 and 2017, indexes including seawater, sediment, seagrass, plankton, zoobenthos and benthic macroalgae was obtained from Liusha bay of Guangdong Province.The results showed that average biomass and total distribution area of Halophila ovalis were 3.74 g/m2 and 26.9 hm2 with patches distribution pattern.Three habitat types were divided by principal component-cluster analysis of seawater parameters, they were "influence area of fish raft and fishing net" (Area Ⅰ), "shellfish farming area" (Area Ⅱ) and "non-cultured area" (Area Ⅲ).Comprehensive scores of principal component showed that Area Ⅰ was most polluted.Comparison between Area Ⅰ and Area Ⅱ showed that biomass of seagrass and zoobenthos, concentration of dissolved oxygen and active phosphate, phytoplankton abundance, salinity in Area Ⅰ were lower than that in Area Ⅱ, biomass of benthic macroalgae, N/P ratio, concentration of ammonia nitrogen, inorganic nitrogen and COD in Area Ⅰ were higher than that in Area Ⅱ.Total distribution area and biomass of seagrass bed in Liusha bay seriously degraded in 2017 when compared with that in 2002.Due to the production mode of fish raft and fishing net, mass multiplication of small shellfishes and low-salinity water derived from river input, seagrass biomass decreased.The relationship between seagrass biomass and other indexes reflected that shellfish culture played a positive role in seagrass survival and strong self-purifying ability of water was beneficial to seagrass growth.Also, this paper put forward the suggestions of protecting seagrass including promoting public consciousness, strengthening natural reserve management and implementating normal monitoring.
-
Keywords:
- Liusha bay /
- seagrass bed /
- principal component-cluster analysis /
- habitat type /
- mariculture
-
海草生活在热带到温带的沿岸海域,在全球的碳、氮、磷循环中扮演着重要角色[1]。海草构成的复杂生境改善了周围的环境[2],为其他生物提供了栖息、庇护和繁殖的场所[3]。海草生长输出的有机质和死后形成的腐殖质,也是复杂的海草食物网的基础[4]。海草床由于其重要的生态价值和功能,和珊瑚礁、红树林一起被称为三大典型海洋生态系统[5]。
广东雷州半岛西南部的流沙湾海域曾分布着我国华南沿海最大的海草床,2002年的面积达900 hm2[6]。最近15 a来,对流沙湾海草床的研究鲜有报导[7]。由于人为因素和自然因素,华南沿岸如海南南部[8]、海南东部[9]、广西合浦[10]和广东沿海[11]等地的大部分海草床都已呈现出退化趋势。流沙湾海草床长期面临着附近居民围网捕鱼、炸鱼电虾和挖贝耙螺等人类活动的威胁[6, 11],加之该海域内海水养殖规模和面积的不断扩大[12],在此背景下进行流沙湾海草床的现状调查研究是十分必要的。为此,笔者于2016~2017年赴流沙湾进行了海草及其环境的调查研究,该研究不仅为流沙湾海草床的后续研究提供基础资料,也为海水养殖环境下的海草床保护和恢复提供参考依据。
1 材料与方法
1.1 站位布设
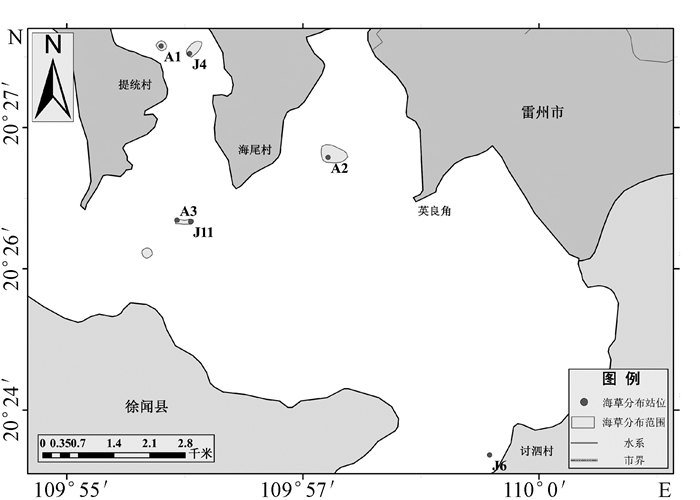
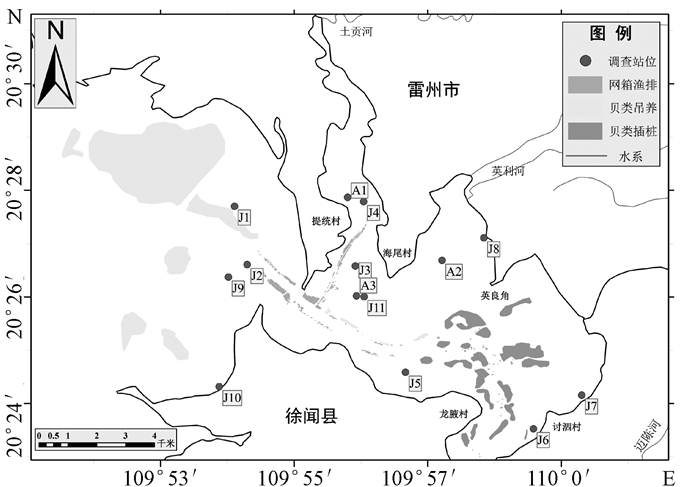
在正式采样之前,笔者通过文献资料[12]、卫星遥感影像和实地调查对流沙湾海水养殖布局进行了大致了解(如图 1所示)。外湾以扇贝吊养为主,在外湾北部有一整片扇贝吊养区。湾口和提统村东侧海域的航道周围,布满了网箱渔排,呈带状分布。内湾常见贝类插桩养殖,多分布在英良角南侧和龙腋村东北侧海域。整个流沙湾还散布有大量的捕鱼网“迷魂阵”,尤以水深较浅的海域居多。调查站位在覆盖所有海草可能分布区域的前提下,兼顾了贝类吊养、插桩区,网箱渔排区和捕鱼网的周围区域。
1.2 采样及分析方法
(1) 海水水质。选取了透明度、pH、盐度、悬浮物、溶解氧、总氮、氨氮、硝酸盐、亚硝酸盐、非离子氨、总磷、活性磷酸盐、CODMn、汞、铜、砷、硫化物和石油类作为监测指标。使用溶氧仪(YSI-556型)现场记录监测海水的溶解氧、盐度和pH,水样经0℃暂存后,迅速带回实验室,按照《海洋监测规范第4部分:海水分析》(GB 17378.4-2007)中的有关规定进行其他参数的检测分析。
(2) 生物参数。选取了浮游植物种类及丰度、浮游动物种类及丰度、底栖动物种类及生物量、海草生物量和大型底栖藻类生物量作为监测指标。使用采水器采集表层水样1 L装入塑料瓶中,加入1%体积分数的鲁格试剂固定作为浮游植物样品。使用装有网口流量计和铅制沉锤的浅水Ⅲ型浮游生物网自底至表进行垂直拖网采集浮游动物,以5%体积分数甲醛溶液固定保存。浮游生物样品迅速带回实验室后静置沉淀24 h,浓缩处理并于倒置显微镜下进行定性和定量分析。在每个有海草分布的站位,进行海草和大型底栖海藻样品的采集。将6.7 cm直径的柱状采样器插入海底10 cm,采集柱状样内所有海草的根、匍匐根、茎、叶片等样品,随后放置50 cm×50 cm大小的不锈钢框,取样框内10 cm海底深度的大型底栖海藻样品。海草和海藻样品离开水面后分别将其放入尼龙网袋,滤掉网袋内泥沙,鉴别后转入塑料袋中封口、编号。样品经-20℃暂存后迅速带回实验室,将剩余泥沙等附着物小心洗涮去除,在60℃恒温条件下持续36~48 h至烘干,称量其干重。单个站位的海草和海藻采样点为3个,采样点相距5~10 m。生物样品的采样和分析方法按照《海草床生态监测技术规程》(HY/T 083-2005)、《海洋调查规范第6部分:海洋生物调查》(GB 12763.6-2007)和《海洋监测规范第6部分:生物体分析》(GB 17378.6-2007)中的有关规定执行。
1.3 数据处理
在每一块海草床边界(主要是拐点位置)设置多个GPS定位点,勾绘海草分布区。使用Google earth和Arcmap 10.2对数据进行空间分析,计算海草床分布面积并作图。使用SPSS 19.0软件将各调查站位的水质参数进行降维处理,提取主成分,并进行聚类分析。使用SPSS 19.0软件对水质参数和生物参数进行Pearson相关性分析和独立样本t检验的差异性分析。
2 结果与讨论
2.1 海草的形态特征和分布
海草全部分布在内湾,共发现了5块海草床和1个海草生长点(如图 2所示),海草种类只有一种,为卵叶喜盐草(Halophila ovalis)(本文海草中文名称参考黄小平等[13])。卵叶喜盐草植株高度为8~12 cm,茎匍匐、纤细,颜色白中带红偏透明,叶片呈卵状椭圆形,较薄且叶脉清晰、凸出,部分叶片有褐色斑纹,叶片长0.6~1 cm,宽度小于0.5 cm,叶柄长0.6~1.2 cm。面积最大的海草床位于英良角西侧、英利河入海口南侧的A2站附近海域,为13.2 hm2。面积次之的海草床位于提统村东侧、土贡河入海口南侧的A1和J4站附近海域,此处退潮时水深仅约1 m,海草床接近片状分布,海草植株较其他区域矮,茎叶较嫩。海尾村西南侧的A3和J11站附近海域海草床呈斑块状分布,海草植株较高,茎叶颜色较深。讨泗村西侧的J6站附近海域水深3~4 m,底泥较多、水体浑浊,未能确定海草床分布面积,此处海草形态特征同海尾村西南侧的类似。卵叶喜盐草的海草床总面积为26.9 hm2,海草生物量变化范围为0.80~6.40 g/m2,均值为3.74 g/m2。
2.2 主成分及聚类分析
2.2.1 主成分分析
海水水质监测数据如表 1所示。通过海水水质参数的主成分分析,得到特征值大于1的前5个主成分(见表 2),其累计贡献率达到82.167%,因此,取前面5个主成分,可反映原始数据的基本信息。
表 1 流沙湾海水水质监测数据Tab. 1 Monitoring data of sea water quality in Liusha bay 表 2 各成分特征值及贡献率Tab. 2 Eigenvector and contribution rate of each components
表 2 各成分特征值及贡献率Tab. 2 Eigenvector and contribution rate of each components
通过回归分析,得出各调查站位在前5个主成分上的得分及调查站位综合得分(见表 3),得分越低污染程度越大。结果显示A2站位污染程度最大,J7站位污染程度最小。6个有海草分布的站位中有4个污染程度排在前4位,依次为A2、A3、J4和A1,另外2个有海草分布的站位J6和J11分别排在第11和12位。
表 3 各调查站点在主成分中的得分Tab. 3 Principal component scores at each sampling sites
2.2.2 聚类分析
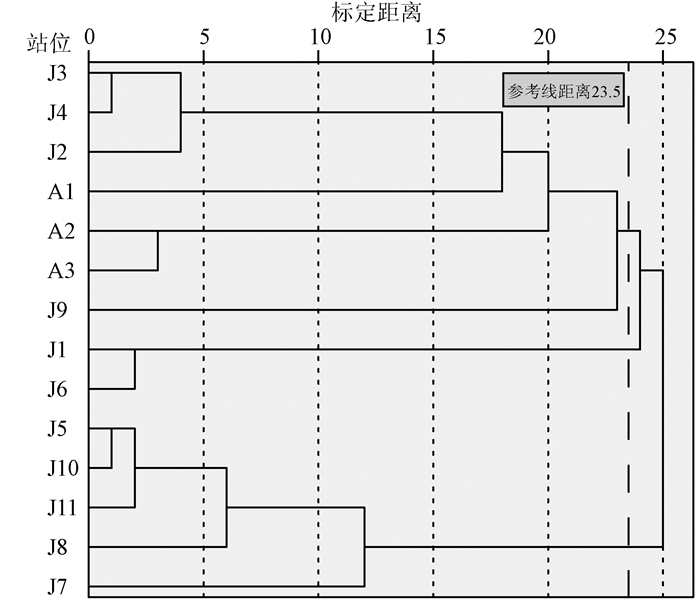
根据实际情况选择Q型聚类方式,使用SPSS19.0软件将表 3中各调查站位在主成分中的得分进行聚类。在分析中采用平方欧式距离,应用系统聚类分析法中的Ward法,采用Z得分对其标准化转换,得到聚类分析树状图(见图 3)。
以距离23.5作为阈值,将各调查站位归类成三种生境类型:渔排和捕鱼网影响区(Ⅰ区)、贝类养殖区(Ⅱ区)和非养殖区(Ⅲ区)。Ⅰ区的渔排是指渔排式网箱,布设在渔排之间航道和渔排周围的调查站位均被划分到了Ⅰ区的渔排影响区里。流沙湾海域捕鱼网“迷魂阵”较常见,许多调查站位都位于捕鱼网附近。由于渔民经常没有及时收获捕鱼网中的鱼虾并清理其中的垃圾,对周围海水环境产生了一定影响,导致捕鱼网影响区和渔排影响区的站位被归为一类。Ⅰ区污染程度综合排名为4.3,是三个生境类型中污染最严重的一个。Ⅱ区中,J1站位为筏式贝类吊养区,J6站位为桩式贝类吊养区,Ⅱ区污染程度综合排名为9.5,比Ⅰ区污染程度轻。Ⅲ区在湾口和在湾内均设有布点,其污染程度综合排名为11.2,污染程度最轻。由于非养殖区海底仍有废弃的蚝桩和捕鱼网,导致该区域未能完全脱离养殖区的影响,分析重点将放在渔排和捕鱼网影响区(Ⅰ区)和贝类养殖区(Ⅱ区)上。各生境类型的主要指标均值如表 4所示。
表 4 各生境类型的主要指标参数Tab. 4 Major index parameters in each habitat type
2.3 流沙湾海草床退化诱因与机理
本研究发现海草床分布面积为26.9 hm2,生物量均值为3.74 g/m2,离2002年海草床分布中心位置最近的A2站生物量为2.32 g/m2。2002年海草床分布面积为900 hm2,生物量均值为25.7 g/m2[6]。以前的海草调查方式是选择当地最大潮差期间的退潮时期,在陆上通过手持GPS仪完成海草分布范围的测绘,而本研究的调查主要通过潜水来完成。如今流沙湾适合海草分布的较浅海域,捕鱼网“迷魂阵”和蚝桩分布密集,加之常年湾内水体能见度较差,这对海草潜水调查,特别是勾绘海草分布范围提出巨大挑战。对比2002年的调查结果,目前流沙湾海草床的分布面积和生物量均低于1/10。虽然调查方式不同可能造成结果的低估,但仍可看出流沙湾海草床已严重退化。
从表 4可知,Ⅱ区和Ⅲ区海草生物量接近,均高于Ⅰ区海草生物量,并超过其2倍,笔者以主成分及聚类分析的结果作为切入点,探讨流沙湾海草的退化原因。
(1) 营养盐竞争处于劣势。Ⅰ区拥有更大的养殖密度,其养殖方式导致水体交换能力更弱,而Ⅱ区和Ⅲ区主要靠近贝类养殖区,其养殖密度相对较小,水体交换能力较好。Ⅰ区网箱产生的大量鱼、虾排泄物及残余饲料等,被细菌分解产生大量氨氮,加之水体交换能力差,导致该区氨氮含量最高。Ⅰ区大量的氨氮消耗更多的溶解氧,使该区溶解氧含量最低。该现象在拓林湾和流沙湾都曾出现过。杜虹等[14]在柘林湾的研究表明养殖区的氨氮含量高于非养殖区,而溶解氧含量较低。谢群等[15]在流沙湾养殖高峰的夏季也观察到了氨氮在四季中最高,且溶解氧含量较低。Ⅰ区大量氨氮在硝化作用下,形成硝酸盐,使该区硝酸盐含量最高。除Ⅰ区A2站位的无机氮超标至《海水质量标准》(GB3097-1997)第三类,其他所有站位的无机氮和活性磷酸盐浓度都符合第二类海水水质标准。三个生境类型的低无机氮和活性磷酸盐浓度都体现了流沙湾的贫营养类型海域特点,与已有的研究结果[16]一致。但也有研究结果[12]表明,流沙湾仅磷酸盐的营养水平属贫营养型。Ⅰ区有较高的生物量和较多科类的大型底栖海藻,是得益于渔排养殖过程产生的高浓度的硝酸盐,而且与浮游植物和海草形成了竞争关系[17-18],并导致了Ⅰ区的浮游植物丰度和海草生物量最低。大型底栖海藻吸收了较多的营养盐,也是流沙湾营养盐含量低、水质长期保持在二类的原因之一[19]。Ⅱ区和的Ⅲ区N/P比值都接近redfield ratio[20]的理论比值16,而Ⅰ区远高于理论比值,达638.26,说明了该区域水体存在磷限制。同时Ⅰ区养殖过程中产生大量有机物形成的微小有机粒子吸附磷酸盐,也会影响海草对其吸收[21],加剧了海草营养盐的竞争劣势。
(2) 生存空间被挤占。Ⅱ区的底栖动物生物量和多样性均高于Ⅰ区。Ⅱ区和Ⅲ区的底栖动物生物量主要由个体较大的帘蛤科和樱蛤科等海洋软相底质底栖贝类贡献,分布较为稀疏。Ⅰ区的底栖动物生物量由占统治地位的汇螺科贡献,虽然其生物量较Ⅱ区低,但个体较小,密密麻麻铺在海底,挤占了海草的生长空间,该现象在广西的合浦海草床已有报导[10]。此外,Ⅰ区的渔排遮挡太阳光线,残饵和粪便覆盖海草及其生长底质,捕鱼网“迷魂阵”打桩时破坏践踏海草,均影响了Ⅰ区的海草生长,这也是华南海草床退化的常见原因[8, 22]。
(3) 近岸河流输入。以盐度为变量,采用和生境类型划分相同的聚类方法对所有站位进行聚类分析,发现Ⅰ区有海草分布的A1、J4和A2站为一类,其他站位为另一类。进一步进行显著性差异分析发现A1、J4和A2站的盐度显著偏低(P < 0.05),该三个站位靠近土贡河或英利河,可知近岸河流输入对盐度产生了影响。该三个站位的海草生物量范围为0.80~2.32 g/m2,远低于其他站位的5.76~6.40 g/m2。有研究表明,卵叶喜盐草生长的最适盐度为30~35[23]或25~35[24]。本研究中受河流输入影响的海草站位盐度范围为22.2~24.8,未受影响的海草站位的盐度范围为26.3~27.3,前者的低盐度更偏离海草的最适盐度,造成了Ⅰ区海草的低生物量。2.4流沙湾海草生物量与其他指标的关系
在对有海草分布和没有海草分布的两组数据进行差异性分析时,发现COD浓度是唯一有显著性差异的参数,有海草分布区域的COD浓度显著高于没有海草分布的区域(P < 0.05)。在贝类养殖的主要季节,流沙湾贝类养殖区的COD浓度明显上升[25]。养殖贝类的排泄物是流沙湾有机物的主要来源,贝类养殖间接地给海草提供了营养物质,并营造了有利于海草生长的生境。差异性分析的结果呈现出流沙湾海草适于COD浓度较高海域生存的现象,表明贝类养殖对海草栖息环境有积极的意义。
对6个有海草分布的站位进行海草生物量与其他指标的相关性分析,结果表明,海草生物量仅与溶解氧浓度和浮游动物多样性指数显著正相关(P < 0.05),与其他指标不相关。何本茂等[26]讨论了铁山港海草生态区中,众多指标与水体净化能力之间的关系,其中包括溶解氧和化学净化能力,以及浮游动物多样性和生物净化能力的正相关关系。由于流沙湾海草区缺少长期、系统的数据,暂时无法对其水体净化能力作深入分析,但是海草生物量与溶解氧浓度和浮游动物多样性指数显著的正相关关系一定程度上反映了流沙湾海草分布区水体自净能力越强,海草生物量越高这一特点。
3 结论
本研究表明,小个体底栖贝类的大量繁殖和河流输入造成的水体盐度波动等自然因素,都会导致海草生物量降低,但流沙湾海草床退化的主要原因是过度使用网箱渔排和捕鱼网,这些生产方式对海草竞争营养盐和占据生存空间都造成了极为不利的影响。贝类养殖对海草的生存和栖息有积极影响,而水体的自净能力会影响海草的生长。
现场调查时发现流沙湾内湾优越的养殖位置均用于渔排养殖和贝类插桩养殖,而在外湾至湾口也有大片的贝类吊养区。此外,见缝插针的“迷魂阵”随处可见,甚至影响到了航道的使用,侵占了流沙湾的公共资源。早在2003年流沙湾就设立了两个县级海洋与渔业自然保护区,分别为3633 hm2的“雷州市海草县级保护区”和300 hm2的“雷州市流沙湾珊瑚礁自然保护区”。流沙湾自然保护区面积占到整个海湾面积的50%以上,但是在现场调查过程中发现保护区中适合养殖的水域都被渔民占用。在沿岸的滩涂,低潮时经常有当地居民在潮间带耙螺挖贝,这种生产方式也极大地影响到了海草的正常生长,导致流沙湾潮间带的海草几乎绝迹。作为最基层的县级自然保护区,其档次、管理水平等方面较国家级、省级和市级自然保护区都存在较大的差距。针对流沙湾海草床的保护工作,笔者提出以下建议:通过宣传画和展览等形式,大力普及海洋生物多样性保护知识,提高当地居民的海草保护意识;加强流沙湾海草床自然保护区的管理,加快海草保护的地方自治立法,规范海水养殖,规制海草床生态系统破坏及污染行为;在海草床主要分布区域选取一条代表性断面开展常态监测,有利于监控流沙湾海草床生态系统状况以及今后的海草恢复工作。
-
[1] FORTES M D.Mangrove and seagrass beds of East Asia:habitat under stress[J].Ambio, 1988, 17(3):207-213.
[2] FONSECA M S.Sediment stabilization by Halophila decipiens in comparison to other seagrasses[J].Estuarine, Coastal and Shelf Science, 1989, 29(5):501-507. doi: 10.1016/0272-7714(89)90083-8
[3] CORLETT H, JONES B.Epiphyte communities on Thalassia testudinum from Grand Cayman, British West Indies:their composition, structure, and contribution to lagoonal sediments[J].Sedimentary Geology, 2007, 194(34):245-262. http://www.sciencedirect.com/science/article/pii/S003707380600176X
[4] DUARTE C M, CHISCANO C L.Seagrass biomass and production:a reassessment[J].Aquatic Botany, 1999, 65(14):159-174. http://d.old.wanfangdata.com.cn/NSTLQK/NSTL_QKJJ02644241/
[5] 宋晖, 汤坤贤, 林河山, 等.红树林、海草床和珊瑚礁三大典型海洋生态系统功能关联性研究及展望[J].海洋开发与管理, 2014, 31(10):88-92. http://d.old.wanfangdata.com.cn/Periodical/hykfygl201410020t [6] 黄小平, 黄良民, 李颖虹, 等.华南沿海主要海草床及其生境威胁[J].科学通报, 200651:114-119. [7] 曾园园.流沙湾喜盐草的生长和生理生化特征[D].: 广东海洋大学, 2015. http://d.wanfangdata.com.cn/Thesis/Y2861360 [8] 陈石泉, 吴钟解, 陈晓慧, 等.海南岛南部海草资源分布现状调查分析[J].海洋学报, 2015, 37(6):106-113. doi: 10.3969/j.issn.0253-4193.2015.06.011 [9] 陈石泉, 王道儒, 吴钟解, 等.海南岛东海岸海草床近10a变化趋势探讨[J].海洋环境科学, 2015, 34(1):48-53. doi: 10.13634/j.cnki.mes20150109 [10] 李颖虹, 黄小平, 许战洲, 等.广西合浦海草床面临的威胁与保护对策[J].海洋环境科学, 2007, 26(6):587-590. doi: 10.3969/j.issn.1007-6336.2007.06.023 [11] 黄小平, 江志坚, 张景平, 等.广东沿海新发现的海草床[J].热带海洋学报, 2010, 29(1):132-135. doi: 10.3969/j.issn.1009-5470.2010.01.020 [12] 罗昭林.流沙湾鱼贝养殖区环境质量评价与浮游生物群落特征分析[D].上海: 上海海洋大学, 2014: 4-18. http://d.wanfangdata.com.cn/Thesis/D599564 [13] 黄小平, 江志坚, 范航清, 等.中国海草的"藻"名更改[J].海洋与湖沼, 2016, 47(1):290-294. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=hyyhz201601035 [14] 杜虹, 黄长江, 董巧香.柘林湾水体溶解氧的分布特征及其与营养盐的关系[J]., 2006, 25(2): 188-193. [15] 谢群, 张瑜斌, 孙省利, 等.流沙湾海域表观耗氧量和营养盐相关性分析[J].环境科学导刊, 2012, 31(2):1-6. doi: 10.3969/j.issn.1673-9655.2012.02.001 [16] 章洁香, 曾久胜, 张瑜斌, 等.流沙湾叶绿素a的时空分布及其与主要环境因子的关系[J].海洋通报, 2010, 29(5):514-520. doi: 10.3969/j.issn.1001-6392.2010.05.007 [17] WEAR D J, SULLIVAN M J, MOORE A D, et al.Effects of water-column enrichment on the production dynamics of three seagrass species and their epiphytic algae[J].Marine Ecology Progress Series, 1999, 179:201-213. doi: 10.3354/meps179201
[18] DELGADO O, RUIZ J, PÉREZ M, et al.Effects of fish farming on seagrass(Posidonia oceanica) in a Mediterranean bay:seagrass decline after organic loading cessation[J].Oceanologica Acta, 1999, 22(1):109-117. doi: 10.1016/S0399-1784(99)80037-1
[19] 张才学, 陈慧妍, 孙省利, 等.流沙湾海草床海域浮游植物的时空分布及其影响因素[J].生态学报, 2012, 32(5):1527-1537. http://d.old.wanfangdata.com.cn/Periodical/stxb201205021 [20] REDFIELD A C, KETCHUM B H, RICHARDS F A.The influence of organisms on the composition of seawater[]1963: 26-77.
[21] HAN Q Y, SOISSONS L M, LIU D Y, et al.Individual and population indicators of Zostera japonica respond quickly to experimental addition of sediment-nutrient and organic matter[J].Marine Pollution Bulletin, 2017, 114(1):201-209. doi: 10.1016/j.marpolbul.2016.08.084
[22] 郭振仁, 黄道建, 黄正光, 等.海南椰林湾海草床调查及其演变研究[J].海洋环境科学, 2009, 28(6):706-709. doi: 10.3969/j.issn.1007-6336.2009.06.026 [23] 杨冉.温度、光照、盐度对喜盐草生长及生理生化特性的影响[D].: 广东海洋大学, 2015: 29-58. http://cdmd.cnki.com.cn/Article/CDMD-10566-1015427418.htm [24] HILLMAN K, MCCOMB A J, WALKER D I.The distribution, biomass and primary production of the seagrass Halophila ovalis in the Swan/Canning Estuary, Western Australia[J].Aquatic Botany, 1995, 51(12):1-54. http://www.sciencedirect.com/science/article/pii/030437709500466D
[25] 王增焕, 柯常亮, 王许诺, 等.流沙湾贝类养殖海域环境质量评价[J].南方水产科学, 2011, 7(3):24-30. doi: 10.3969/j.issn.2095-0780.2011.03.005 [26] 何本茂, 韦蔓新, 李智.铁山港海草生态区水体自净能力与水、生、化之间的关系[J].海洋环境科学, 2012(5):662-666. http://d.old.wanfangdata.com.cn/Periodical/hyhjkx201205009 -
期刊类型引用(7)
1. 张立,郭康丽,党二莎,索安宁,何伟宏. 贝克喜盐草在珠海唐家湾的分布及其生长特征. 海洋湖沼通报(中英文). 2025(02): 1-8 .  百度学术
百度学术
2. 钟超,廖亚琴,刘伟杰,隋昊志,陈清华. 广东沿海海草床的现状、面临的威胁与保护建议. 生物多样性. 2024(02): 118-135 . 百度学术
3. 周慧芝. 广东省红树林和海草床存在的问题与保护建议. 棉花科学. 2024(08): 143-145 . 百度学术
4. 黄盛,胡慧娜,石荣贵,杨熙,余威,邓伟. 广东流沙湾海草床生态系统服务价值评估. 湿地科学与管理. 2023(03): 33-37 . 百度学术
5. 彭宇航,张才学,孙省利,郑金容,刘耀谦,姚昕. 雷州半岛沿岸海域浮游桡足类的分布特征及其影响因素. 海洋与湖沼. 2022(02): 375-383 . 百度学术
6. 邓筱凡,张宏瑜,吴忠迅,李文涛,张沛东. 荣成马山里海域海草床分布现状及其生态特征. 海洋学报. 2022(08): 97-109 . 百度学术
7. 徐步欣,张剑,郎尚昆,陈石泉,吴钟解,王道儒. 海南文昌沿岸海草床的现状及其退化因素分析. 应用海洋学学报. 2022(04): 614-624 . 百度学术
其他类型引用(2)
 下载:
下载:
计量
- 文章访问数: 10237
- HTML全文浏览量: 10366
- PDF下载量: 56
- 被引次数: 9
